最近,华南师范大学生物光子学研究院邢达教授研究团队在Cell Press细胞出版社旗下期刊Stem Cell Reports上发表了一篇题为“Photobiomodulation therapy for hair regeneration: A synergetic activation of β-CATENIN in hair follicle stem cells by ROS and paracrine WNTs”的文章,研究团队在探讨光生物调节疗法(PBMT)促进毛发再生的机制领域取得重要研究进展。该团队研究表明,毛囊干细胞产生的ROS和真皮分泌的WNTs协同激活GSK-3β/β-catenin信号,进而促进毛发再生。拒绝“秃”如其来,Wnt/β-catenin可谓功不可没。今天,就让小编带您来揭秘Wnt/β-catenin有哪些王牌武将和混世之道?
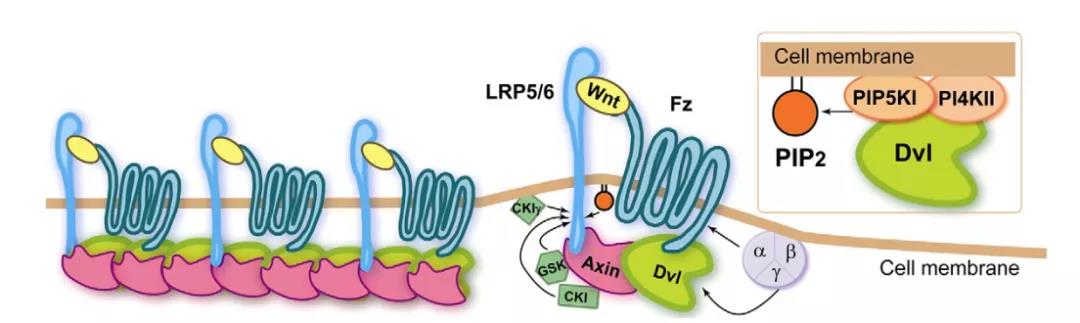
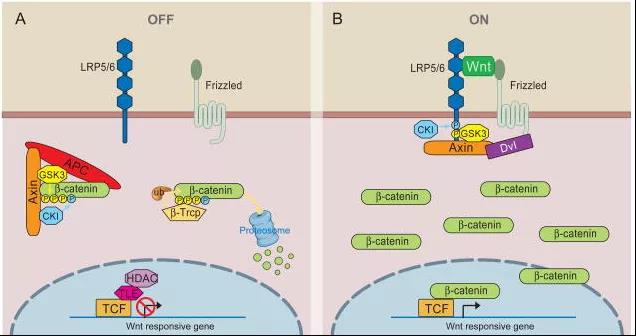
1982年,科学家Nusse与Harold Varmus首次从小鼠乳腺癌模型中鉴定出Wnt基因,因为可以激活乳腺癌相关基因的插入(integration,简称Int),并使正常果蝇表现为无翅(Wingless,简称Wg),将Wg和Int结合便称为Wnt基因。Wnt是一组富含半胱氨酸的分泌性糖蛋白,作为局域性信号分子,广泛存在于动物多种组织中。β-catenin是果蝇Arm蛋白的同源物,其在胞质内降解及在核内积累,可激活一系列靶基因的转录导致肿瘤发生,在Wnt信号通路中起到核心作用。 Wnt/β-catenin作为Wnt经典信号通路途径,极为保守。从低等动物(如线虫)到高等动物(如哺乳类)都具有高度的同源性,可谓“行业老炮”。从崭露头角到行走江湖。当然,Wnt/β-catenin麾下也不乏王牌武将,号称“八大金刚”: ①Wnt家族分泌蛋白 在Wnt信号通路中充当配体,激活信号通路,没有Wnt,β-连环蛋白不会在细胞质中积累。 ②β-连环蛋白(β-catenin) 在胞质中积累继而向细胞核转位,被认为是Wnt信号通路被激活的标志,同时也参与细胞间的黏附。 ③膜受体卷曲蛋白(Fzd/Frz) 分泌型糖蛋白Wnt的细胞膜上受体,为7次跨膜蛋白,结构类似于G蛋白偶联型受体。N端有一个富含半胱氨酸的CRD结构域,能与Wnt结合。 ④蓬松蛋白(Dsh/Dvl) 经过Wnt蛋白处理后,Dsh/Dvl开始过度磷酸化,并抑制糖原合成激酶,进而促进β-catenin稳定。 ⑤糖原合成激酶3(GSK3) 是一种丝氨酸/苏氨酸蛋白激酶。在没有Wnt信号时,GSK3能将磷酸基团加到β-Catenin N端的丝氨酸/苏氨酸残基上,促进了β-Catenin的泛素化及降解。 ⑥支架蛋白(Axin) 作为一种支架蛋白,能与APC、GSK3β、CK1等形成β-Catenin降解复合物。 ⑦结肠腺瘤性息肉病基因(APC) 在Wnt信号不存在的情况下,GSK-3β使APC/β-Catenin/Axin复合体的三个成员磷酸化作为泛素提供识别位点,促使β-Catenin降解。 ⑧T细胞因子/淋巴增强因子(TCF/LEF) 是一类具有双向调节功能的转录因子,可与Groucho结合可以抑制基因转录,而与β-Catenin结合则促进下游靶基因的转录。 另外,也有众多“太保”如辅助受体LRP5/6、酪蛋白激酶(CK1)等,这些成员彼此之间合纵连横,助力Wnt/β-catenin运筹帷幄,决胜千里。 图1 Wnt/β-catenin信号通路潜在的治疗靶点[1] 一、当细胞缺乏Wnt信号时 二、Wnt信号水平较高时 图2 Wnt/β-catenin信号通路图[2]