攻击性被定义为在资源竞争或社会等级背景下发生的行为表现和身体攻击。在多种社会物种中都观察到了攻击性行为,例如海豚、黑猩猩和不同种类的鸟类。在整个动物界中,经常可以观察到同一物种成员之间的种内攻击和不同物种成员之间的种间攻击。这两种类型的攻击行为通常源于对食物的竞争、空间竞争和生存竞争。Volker和Herzing在《动物行为与认知》上发表了一篇关于海豚之间攻击性互动的文章[1],为我们介绍了野生海豚的攻击行为。
同域物种
在巴哈马群岛,大西洋斑点海豚(Stenella frontalis)和宽吻海豚(Tursiops truncates)是同域物种,这意味着它们的亲缘关系非常密切,同时也栖息在同一地理区域。这些物种会一起旅行和觅食,但它们也会进行攻击性互动,攻击行为可能包括头对头的姿势、张开嘴展示、拍打尾巴和下巴、身体冲锋、展示圆形气泡等。
图1巴哈马研究区域
种内和种间的攻击性相互作用
在整个动物王国中,动物都会表达对他人的攻击性。这可以在同一物种的成员之间(种内)或不同物种的成员之间(种间)。这种竞争通常源于对食物的竞争或对配偶的竞争,特别是为了防止两个物种同域生活时的杂交。
在黑猩猩和鸟类中都观察到,根据对手的不同,它们会使用不同的攻击策略。而群体内造成的伤害不如群体间造成的伤害严重。本研究的目的就是调查大西洋斑点海豚群体在种间和种内攻击过程中使用的三种攻击行为:接触行为、追逐行为和展示行为类型。
研究方法
(一)定义攻击性、种内和种间
攻击性被定义为在一次相遇过程中发生的多种多样行为表现和身体攻击。种内攻击行为被定义为针对同一物种个体的攻击行为,而种间攻击行为则针对另一个物种的个体(在本例中为宽吻海豚)。种内或种间攻击行为被定义为在一段时间内发生的多种种内或种间攻击行为。
(二)来自野生海豚项目数据库的视频选择
在本研究中,从野生海豚计划长期数据库中选择了 1991 年至 2004 年期间的水下遭遇档案资料。提取了22个视频,其中 11个是种间攻击,11个是种内攻击。视频记录在任何给定的遭遇中都是连续的,时间范围从5分35秒到21分30秒不等。行为样本的较小片段,具有足够的高分辨率,画面清晰,进而进行组提取分析。
(三)使用行为观察记录分析系统
行为观察记录分析系统用于集成视频进行行为编码和评分。行为被编码到系统中,以对种内和种间的攻击活动进行评分。研究者开发了行为图,行为被分为三类:
展示:拱起、产生气泡、张开嘴、拍下巴等。
追逐:追逐或跟随另一只海豚
接触:猛冲、按住、咬、拍尾巴等。
完整的行为图如下表1所示。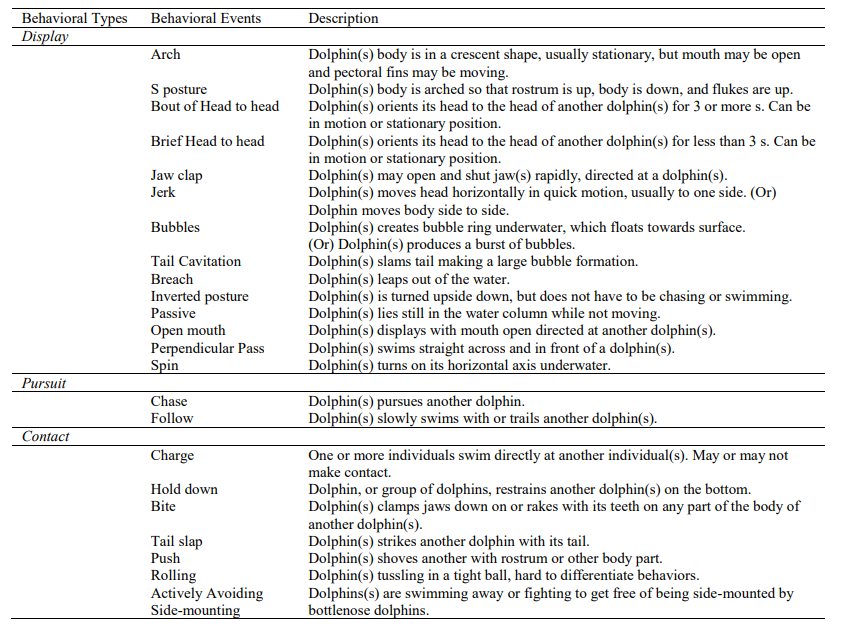
表1 行为类型和行为事件表
行为观察记录分析系统是一款专业用于收集、分析、展示观察数据且操作简便的行为事件记录软件。系统可以支持和引导研究项目的整个工作流程,帮助完成数据采集、管理、筛选、可视化和分析工作。系统可以定量编码行为并在时间轴上可视化呈现,轻松整合视频和生理数据,计算统计数据并评估数据可靠性。同时,系统能够连接不同的硬件进行使用,与数据采集系统的接口或同步其它程序和系统。
结果
1991年至2004年间,总共记录了 1,550 次遭遇事件。种内遭遇(仅限斑点海豚)占总数的75.78%,其中 9.86% 的遭遇中观察到种内攻击行为。种间遭遇(斑点海豚和宽吻海豚)占同一时间段内遭遇总数的 14.7%,其中 45.2% 发生了种间攻击。
(一)群体规模分析
群体规模对种间攻击(U = 10,p = .361)或种内攻击(U = 10,p = .683)的遭遇行为率没有显著影响。 因此,11 个种内遭遇和 11 个种间遭遇可以合并并作为两个单独的组进行分析。
(二)比较种内和种间攻击
所有行为事件的每分钟平均速率
当检查所有行为事件的遭遇行为率时,种间和种内攻击之间的平均行为率没有显著差异(U = 56.000,p = .797)(Minter = 8.12,Mintra = 7.60,图 2)。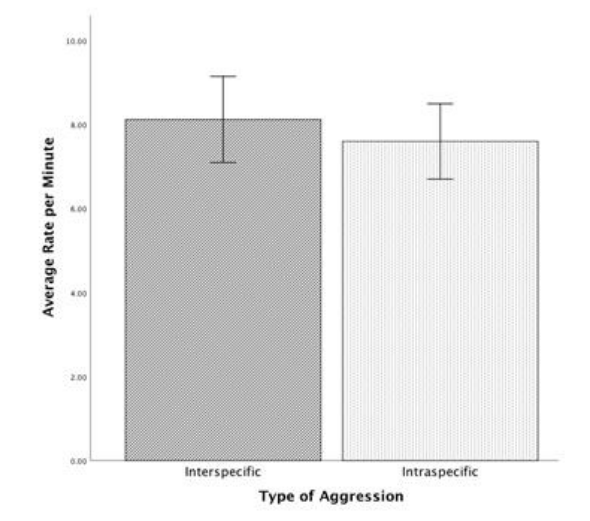
图2
种内和种间攻击遭遇期间所有行为事件组合的每分钟平均速率
行为类型
在种间和种内攻击期间,这三种行为类型以相同的频率模式使用。“展示”的每分钟平均速率(rpm) 最高(Minter = 4.26,Mintra = 5.49),其次是“接触”(Minter = 2.45,Mintra = 1.78),而“追逐”的每分钟平均速率最低(Minter = 1.42,Mintra = 0.33)。
当比较种间攻击和种内攻击之间的行为类型的平均速率时,Mann-Whitney U 测试表明,在种间攻击期间,“追逐”的使用明显多于在种内攻击期间的使用(U = 9,p = .001,图 3) 。 “展示”(U = 40,p = .178)或“接触”(U = 37,p = .123)行为类型的使用没有显著差异(图3)。
对于种间攻击性,Friedman检验显著。每分钟使用的“展示”和“接触”中位数均显著高于“追逐”(“展示”与“追逐”,p = .001;“接触”与“追逐”,p = .012)。然而,“展示”和“接触”之间没有显著差异(p = .065,图 3)。
对于种内攻击性,Friedman 检验也显著。所有三种行为类型均引发显著不同的中位数。“展示”每分钟使用率中值显著高于“接触” (p = .001) 和“追逐”(p = .001),并且“接触”的每分钟使用率中值显著高于“追逐”(p = .001,图 3)。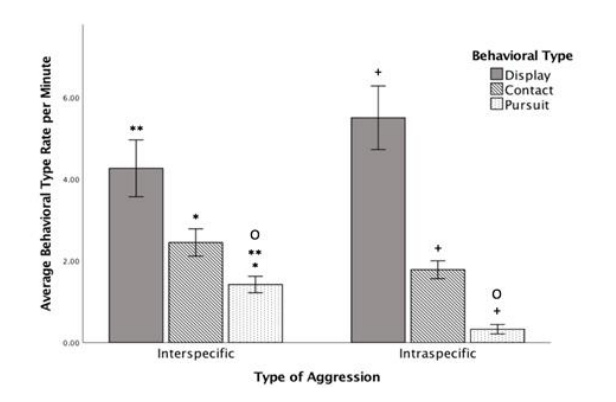
图3
每种行为类型在种间攻击和种内攻击中的平均每分钟使用率
个别行为事件
以下个别行为事件在种内和种间攻击期间每分钟的平均速率最高:猛冲(接触)、短暂的头对头(展示)和张开嘴(展示)(图 4)。在总共 23 个行为事件中:三个仅在种间攻击期间观察到(主动避免侧挂、被动、尾部气穴),而两个仅在种内攻击期间观察到(S姿态、旋转,表2)。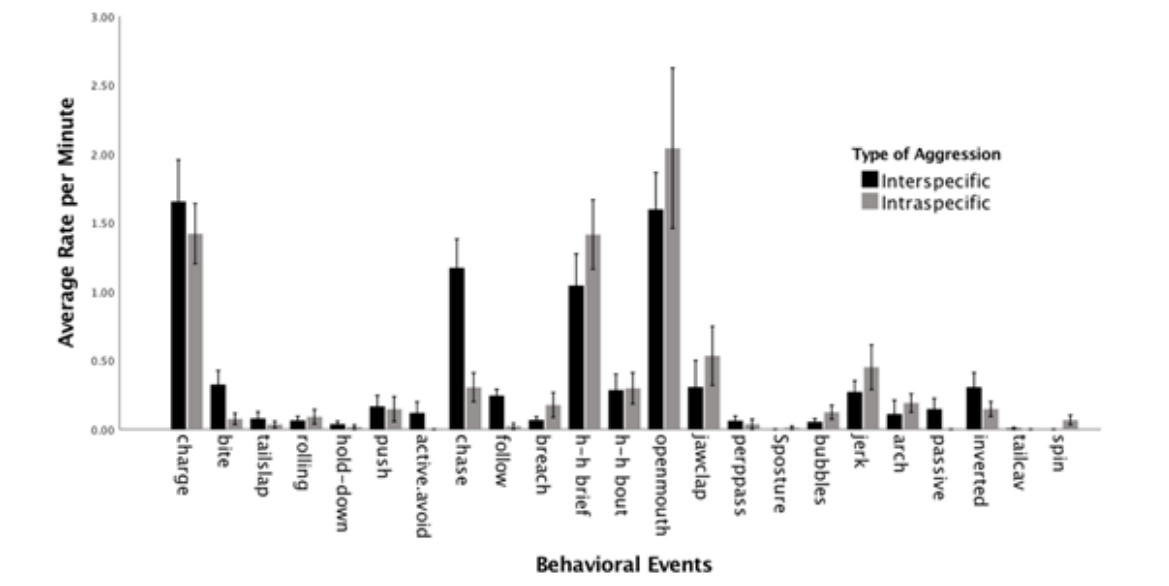
图4
在种间和种内攻击期间每分钟观察到每个行为事件的平均速率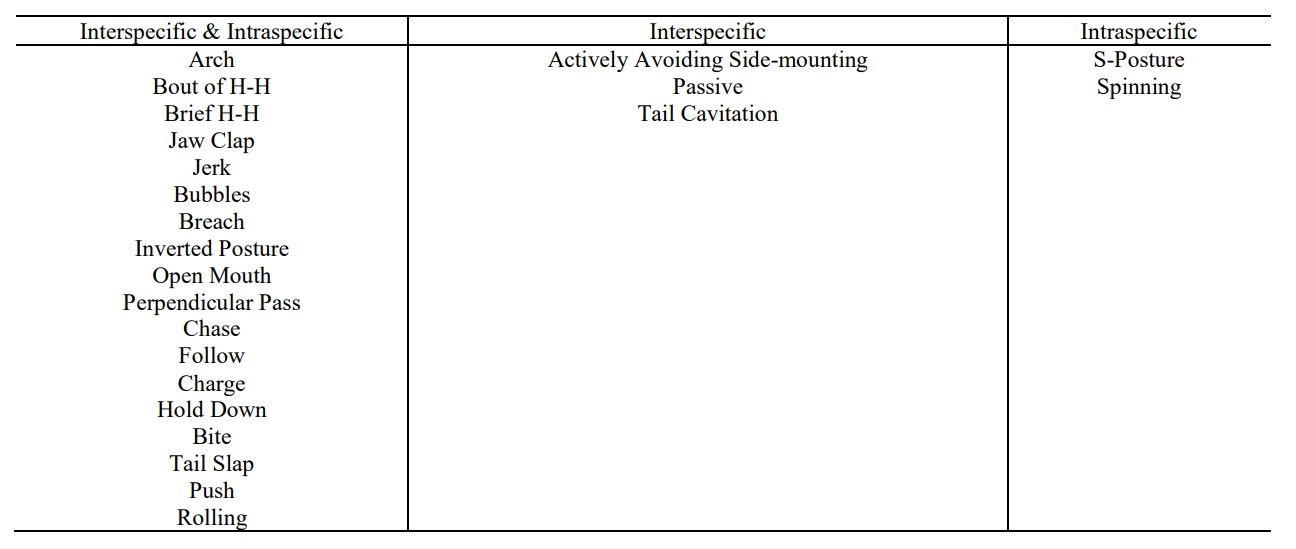
表2
在两种类型的攻击、仅种间攻击或仅种内攻击期间观察到的行为事件
发现
研究发现总体行为率没有差异,这意味着这些海豚在任何一种攻击行为中都没有使用更具攻击性的事件。每种攻击类型都有独特的行为事件,例如尾部气穴(种间)和旋转(种内)。
(一)种内和种间攻击行为事件
在种间和种内攻击期间,斑点海豚使用的行为事件的总体平均行为速率(rpm)没有显著差异:在任何一种类型的攻击期间,它们都没有使用明显更具攻击性的行为事件。
斑点海豚群体在攻击过程中总共使用了 23 种不同的个体行为事件。在种间和种内攻击期间使用了十八个行为事件。其他五种行为事件仅记录在一种攻击类型中(表 2)。在这两种类型的攻击中,使用最多的是相同的三种行为事件:猛冲、短暂的头对头和张开嘴。除其他外,这三种行为也在宽吻海豚的攻击行为中曾被观察到。这两个物种之间存在和使用类似的攻击行为并不太令人惊讶。宽吻海豚和斑点海豚具有相同的流线型身体结构,限制了它们扭曲和操纵身体的方式,导致行为相似,遗传学和类似的社会结构也可以解释攻击行为过程中的相似性。
在大西洋斑点海豚的攻击行为期间,总共观察到了展示行为类型中的 14 种不同的展示行为事件。与接触和追求相比,展示行为类型的个体行为事件数量也最多,平均使用次数也最多。 这并非没有道理,因为个人通常会先展示来威慑敌人,以避免浪费体力,同时也减少受伤的机会。大量的展示行为对于一个物种来说可能是有利的。“分级信号假说”可能为该群体中各种展示行为提供可能的解释。分级信号是强度增加的警告,如果敌人不退缩,则意味着更严重的后果。根据情况或对手的不同,斑点海豚在对抗过程中似乎有大量的展示行为可供选择。
(二)种内和种间攻击行为类型
斑点海豚群体在种内和种间攻击过程中都以相同的总体模式使用三种行为类型:展示行为的每分钟平均速率 (rpm) 最高,其次是接触行为,而追逐行为类型的平均速率最低。从进化的角度来看,展示行为被认为比接触和追逐行为消耗的能量更少,风险也更小。这可能解释了为什么斑点海豚最常利用它们的展示行为,正如前文所述,许多物种都使用威胁展示来传达意图,并且可用于在冲突升级为全面的物理攻击之前解决冲突。
当分别检查每种攻击类型的行为类型时,也发现了一些差异。在种内攻击期间,斑点海豚使用展示型的次数明显多于接触型,但在种间攻击期间,这两种类型的使用并没有显著差异。 宽吻海豚比斑点海豚体型更大,因此,在种间冲突中,数量优势对斑点海豚也起着至关重要的作用。 Herzing 和 Johnson(1997)曾研究表明,斑点海豚与宽吻海豚的比例至少需要六比一,才能使攻击性向有利于它们的方向发展[2]。在种内攻击过程中,斑点海豚可能会使用比展示行为少得多的接触行为,以降低同伴受伤的风险,在其他攻击性遭遇中,它们可能需要这些同伴来帮助对抗更大的宽吻海豚。
另外,在种间攻击过程中同样会使用能量消耗更高的行为类型:“追逐”。如上所述,接触和追逐行为可能比展示行为更耗费能量且风险更大。并且,当斑点海豚与宽吻海豚搏斗时,接触的使用并不明显少于展示,这表明斑点海豚在种间攻击过程中也会使用较多的能量。当个体将更多精力投入到攻击性互动中时,就像斑点海豚在与宽吻海豚发生攻击性遭遇时所做的那样,表明宝贵的资源受到威胁。种间攻击的原因可能包括对食物、领地或交配机会的竞争,正如其他哺乳动物物种中所发现的那样。
结论
本文研究发现,两种海豚会根据体型使用不同的策略:在种内(同一物种)攻击过程中,斑点海豚使用展示策略的次数比使用接触策略的次数多,而在种间(不同物种)攻击时,这两类海豚的使用次数并不比另一类多。斑点海豚比宽吻海豚小,因此需要数量上的优势才能使攻击性朝着对它们有利的方向发展。这些都表明,通过根据对手改变它们的攻击行为,这些海豚确实具有高度的行为可塑性。
行为可塑性让海豚可以根据环境中变化的刺激来学习和改变自己的行为,包括物种重叠的可能性。当群体因气候变化而被迫进入新的栖息地,并不得不适应才能生存时,这一点发挥了重要作用。
参考文献
1.Volker, C. L.; Herzing, D. L. (2021). Aggressive Behaviors of Adult Male Atlantic Spotted Dolphins: Making Signals Count during Intraspecific and Interspecific Conflicts. Anim. Behav. Cogn., 8 (1), 35–51
2.Herzing, D. L., & Johnson, C. M. (1997). Interspecific interactions between Atlantic spotted dolphins (Stenella frontalis) and bottlenose dolphins (Tursiops truncatus) in the Bahamas, 1985-1995. AquaticMammals, 23,85–99.
关注诺达思公众号,联系我们获取更多产品信息及学术文章!
测量野生海豚的攻击行为
作者:诺达思(北京)信息技术有限责任公司 2024-01-19T00:00 (访问量:27190)
诺达思(北京)信息技术有限责任公司 商家主页
地 址: 北京市朝阳区广顺北大街33号院6号楼福泰中心507室
联系人: 诺达思
电 话: 010-84851148/18501168738
传 真:
Email:info-china@noldus.com;
相关咨询
如何用彩色迷宫破解阿尔茨海默症奥秘? (2025-03-20T00:00 浏览数:3930)
基于行为观察的居民生活空间设计洞察 (2025-03-14T00:00 浏览数:5940)
为什么要在行为研究中测量皮肤电反应? (2025-03-14T00:00 浏览数:6238)
田鼠的跨代智慧:捕食者气味如何影响后代行为? (2025-03-07T00:00 浏览数:7236)
斑马鱼暴露于大麻酚研究 (2025-02-28T00:00 浏览数:6880)
线上心理咨询中“好咨询师”的面部表情 (2025-02-25T00:00 浏览数:9116)
用于治疗肩袖损伤的缝合锚 (2025-02-21T00:00 浏览数:8614)
行为观察与模拟技术帮助优化手术室设计 (2025-02-19T00:00 浏览数:10876)
在三维轨迹中分析蚊子寻找宿主行为 (2025-02-14T00:00 浏览数:10849)
城市光环境对情绪影响的多模态研究 (2024-12-19T00:00 浏览数:21882)