【干货】拯救你受伤的DNA-NHEJ与HR
生命极其脆弱,我们每天在电子辐射、紫外线、雾霾等等各种外部环境及细胞代谢产物等内源因素影响下,我们生命的核心-DNA都会受到不同程度的损伤,其中DNA双链断裂(DSBs,Double strand breaks)是损伤中最为严重的一种,然而生命却又极其强大,我们无时无刻不在受伤,也无时无刻不在自我修复。那么我们的身体里是用什么样的机制修复DNA损伤呢?且听我们慢慢道来~
对于DSBs损伤,主要有非同源末端链接(Non-homologous end joining,NHEJ)和同源重组(Homologous recombination,HR)两种,单链退火修复一般发生在串联同源DNA序列中,研究较少[1](图1)。下面小编就给大家具体介绍一下日常体内是如何通过这两种方式修复拯救我们受伤的DNA的。
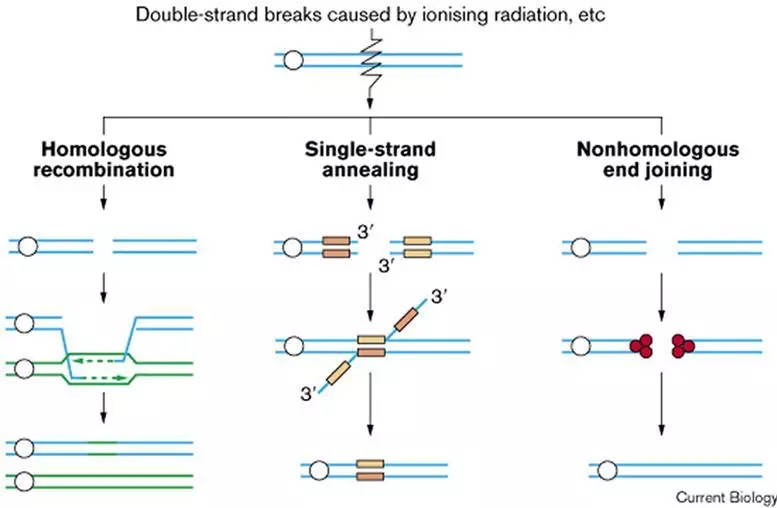
图1. DSBs 修复途径
Alternative pathways of DNA double-strand break repair. Homologous recombination is the preferred route in yeast. It involves invasion of the broken DNA strands into a homologous DNA duplex molecule. This process requires Rad52 (a DNA end-binding protein), Rad51 (which forms filaments along the unwound DNA strands), DNA polymerases and other less well-characterized gene products. The DNA ends are ligated by DNA ligase I and the interwound DNA strands are separated, probably by another protein complex, with no loss of genetic information. Only one of many possible recombination products is shown here. Single-strand annealing takes place between two homologous DNA sequences in tandem (yellow and orange boxes) by a less well-studied mechanism. It also requires Rad52, and extensive degradation of the two unannealed strands results in considerable loss of genetic material. Nonhomologous end joining rejoins the two broken ends directly. It requires the DNA end-binding protein Ku which, in mammalian cells, forms a complex with DNA-PKcs(red). Other steps in this process involve the Rad50–Mre11—Nbs1 complex in mammals (which may also be involved in homologous recombination, at least in yeast) and the XRCC4—DNA ligase IV complex. Few, or no, bases are missing from the products of nonhomologous end joining.
非同源末端链接-NHEJ
NHEJ是一种简单粗暴高效的修复方式,断裂了?管它三七二十一,赶快连起来!就是这个暴脾气!

NHEJ: 我这暴脾气![6]
它就是通过DNA连接酶将DSBs末端直接链接的一种修复过程,不依赖于同源DNA序列。Ku蛋白(Ku70/Ku80)复合物识别结合到DSBs末端,Ku-DNA复合物招募DNA依赖蛋白激酶催化亚基(DNA-PKcs)激活其激酶活性,将自身磷酸化启动NHEJ通路,吸引重组酶Artemis加入处理DNA末端,然后召集XRCC4- DNAligase4 –XLF复合物促使DNA末端进行连接[2] 。
NHEJ这种暴脾气虽然迅速高效,但是会有一系列副作用,就是可能造成一些序列缺失,同时也会造成一些片段的插入,即所谓不够精确。
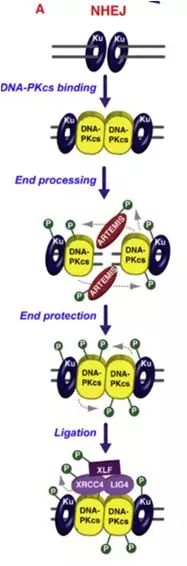
图2. NHEJ
Rapid association of Ku to DSBs promotes NHEJ by recruiting DNA-PKcs. Sequential phosphorylation events on multiple DNA-PKcs amino acid clusters favors the initial processing of DNA ends by ARTEMIS, followed by DNA-PKcs-dependent protection of DNA ends required for DNA ligation.
同源重组-HR
HR是另一种处女座式追求怀旧与完美的修复方式:我要找到我自己!一个都不能多!一个都不能少!

HR: 我就是这么完美![7]
HR需要以未受伤的姐妹染色单体的同源序列作为其修复的模板。MRN复合物识别DSBs,结合到DNA末端,修复第一步就是要将DNA末端进行修剪,MRN复合物和转录因子CtIP(CtBP-interacting protein)促进DNA末端切割过程,造成5’末端DNA降解,产生3’单链DNA(ssDNA),3’ ssDNA被复制蛋白A(Replication protein A,RPA)包被,使其免受核酸酶的降解,去除二级结构;然后由BRCA2蛋白介导,RPA被重组酶RAD51替换,形成核蛋白丝寻找姐妹染色单体上的同源序列,RAD51蛋白介导侵入DNA双链模板,与同源DNA序列配对形成D-Loop结构,D-Loop延伸或与另一个末端连接,完成修复过程[3] 。

图3. HR
The MRN-CtIP-complex starts resection on the breaks to generate single stranded DNA (ssDNA). After resection the break can no longer be repaired byNHEJ. The ssDNA is first coated by RPA, which is subsequently replaced by Rad51 with the help of BRCA2. These Rad51 nucleoprotein filaments mediate strand invasion on the homologous template. Extension of the D-loop and capture of the second end lead to repair.
HR强迫症式的修复,虽然速度较慢,效率较低,但是精准,可以使基因组得到完美修复,完好如初!
经过这一番介绍,大家对我们的DNA双链修复小伙伴应该有了初步印象了吧?
简言之,他们的特点就是:NHEJ的修复是快速非精确的,过程中可能会随机的引入和去除几个碱基,整个细胞周期都有发生,在G1期及S期发挥着重要作用;HR的修复是复杂而精确的,但是需要同源序列模板,只能发生在细胞G2/S期。针对不同DSBs损伤类型机体会选择性的启动NHEJ或HR修复机制,拯救已经受伤的DNA。
说到这里,大家是不是对DNA修复机制加深了一点了解呢,只有掌握学习了才能更好的应用。
学以致用
随着科学家们对这两种修复方式的深入研究和解读,这一修复机制被逐渐应用于基因编辑技术,用于定向改造基因。研究者采用核酸酶技术使DNA双链有目的的产生断裂,利用这一机制调控目的基因表达或引入选择性标记。
没有模板的情况下,利用NHEJ修复,随机插入或缺失碱基造成基因失活;有模板存在的情况下,可通过同源重组的方式在剪切位点引入目的修饰基因,实现基因的定点敲进[4] (图4)。
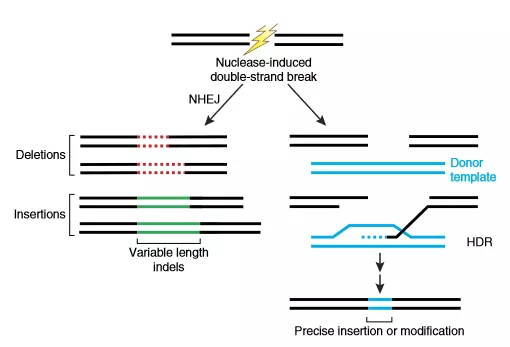
图4. Nuclease-induced genome editing
Nuclease-induced double strand breaks (DSBs) can be repaired by nonhomologous end joining (NHEJ) or homology-directed repair (HDR) pathways. Imprecise NHEJ mediated repair can produce insertion and/or deletion mutations of variable length at the site of the DSB. HDR-mediated repair can introduce precise point mutations or insertions from a single-stranded or double stranded DNA donor template.
以CRISPR/Cas9技术为例,如图5a,Cas9蛋白剪切DNA双链形成DSBs,利用NHEJ修复机制,随机缺失或插入碱基造成移码突变,目的基因实现敲除。图5b,剪切同时引入一段外源基因,NHEJ会将其接入DSBs位点,实现基因插入。图5c,两端分别设置剪切位点,中间片段游离,两端通过NHEJ机制连接形成大片段基因敲除[4] 。
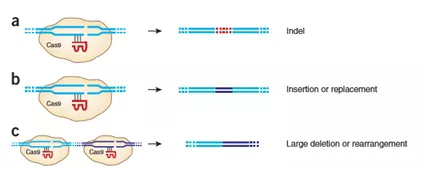
图5. (a,b) gRNA-directed Cas9 nuclease can induce indel mutations (a) or specific sequence replacement or insertion (b). (c) Pairs of gRNA-directed Cas9 nucleases can stimulate large deletions or genomic rearrangements (e.g., inversions or translocations).
剪切的同时加入带外源基因及同源序列,HR机制作用下可实现基因定点精确的敲进和替换。CRISPR/Cas9 介导产生NHEJ的效率远远高于HR[5],怎么能让完美主义的HR小伙伴发挥更大的作用,提高我们的基因敲进效率呢?
为了实现高效精确的基因定点敲进,百奥赛图自主研发的EGE(Extreme Genome Editing System)系统,将同源重组效率提高了近20倍(图6),很大程度的提供了更多精确编辑基因的可能!

图6. 百奥赛图自主研发的EGE(Extreme Genome Editing System)系统
百奥赛图具有多年制备基因敲除敲进大小鼠的经验,专业的技术团队,需要制备的老师请通过下方联系方式联系我们哦。
参考文献:
1.Featherstone, C., & Jackson, S. P. (1999). DNA double-strand break repair. Current Biology, 9(20), R759–R761.
2.Ciccia, A., & Elledge, S. J. (2010). The DNA Damage Response: Making It Safe to Play with Knives. Molecular Cell, 40(2), 179–204.
3.Brandsma, I., & Gent, D. C. (2012). Pathway choice in DNA double strand break repair: observations of a balancing act. Genome Integrity, 3(1), 9.
4.Sander, J. D., & Joung, J. K. (2014). CRISPR-Cas systems for editing, regulating and targeting genomes. Nature Biotechnology, 32(4), 347–355.
5.Ma, Y., Chen, W., Zhang, X., Yu, L., Dong, W., Pan, S., … Zhang, L. (2016). Increasing the efficiency of CRISPR/Cas9-mediated precise genome editing in rats by inhibiting NHEJ and using Cas9 protein. RNA Biology, 13(7), 605–612.
6.https://www.jiuwa.net/photo/43172/
7.http://www.yixiaoxi.com/renwuwanmeiyikewanshan.html